GROWTH AND LIFE HISTORY

Digitization of rare and endangered strepsirrhine primates from the Duke Lemur Center
For the past three years, ongoing efforts at Duke University have resulted in microCT scanning of more than 100 strepsirrhine primate cadavers (17 species) from the Duke Lemur Center. To date, more than 500 scans have been generated, including specimen overviews and zooms of complex anatomical regions (e.g., cranium, hands, feet). Scans are publicly available through the digital repository Morphosource.org. As captive (but free-ranging) individuals, these specimens have a wealth of associated information that is largely unavailable for wild populations, including detailed life history data, longitudinal body mass, and records of use in experiments. This collection is also exceptional in representation of age categories that are uncommon in museum collections, and maintains associations in indistinct anatomical regions (e.g., caudal vertebrae, manual and pedal phalanges). Currently, these specimens are stored frozen and are available for a fee to research projects that require destructive sampling. This digital collection maximizes the information obtained from rare and endangered animals without degradation of the original specimens.
(Gif courtesy of Justin Gladman).
For the past three years, ongoing efforts at Duke University have resulted in microCT scanning of more than 100 strepsirrhine primate cadavers (17 species) from the Duke Lemur Center. To date, more than 500 scans have been generated, including specimen overviews and zooms of complex anatomical regions (e.g., cranium, hands, feet). Scans are publicly available through the digital repository Morphosource.org. As captive (but free-ranging) individuals, these specimens have a wealth of associated information that is largely unavailable for wild populations, including detailed life history data, longitudinal body mass, and records of use in experiments. This collection is also exceptional in representation of age categories that are uncommon in museum collections, and maintains associations in indistinct anatomical regions (e.g., caudal vertebrae, manual and pedal phalanges). Currently, these specimens are stored frozen and are available for a fee to research projects that require destructive sampling. This digital collection maximizes the information obtained from rare and endangered animals without degradation of the original specimens.
(Gif courtesy of Justin Gladman).
| Yapuncich et al 2019 - Digital collection of DLC primates |
ALLOMETRY
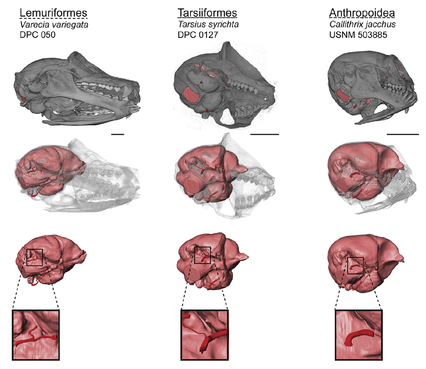
Cranial blood supply in euarchontans
Primates differ from other mammals in having bony canals that enclose the branches of the internal carotid artery (ICA) as they pass through the middle ear. The presence and relative size of these canals varies among major primate clades. As a result, differences in the anatomy of the canals for the promontorial and stapedial branches of the ICA have been cited as evidence of haplorhine or strepsirrhine affinities among early fossil euprimates. Here we use microCT to compile the largest quantitative dataset on ICA canal sizes. The data suggest greater variation of the ICA canals within some groups than has been previously appreciated. In species where the promontorial artery supplies the brain with blood, the size of the promontorial canal is significantly correlated with endocranial volume (ECV). In species with alternate routes of encephalic blood supply, the promontorial canal is highly reduced relative to ECV, and correlated with both ECV and cranium size. Ancestral state reconstructions incorporating data from fossils suggest that the last common ancestor of primates had promontorial and stapedial canals that were similar in size and large relative to ECV. This condition is exhibited by treeshrews and most early fossil euprimates, while extant primates exhibit reduction in one canal or another.
Boyer DM, Kirk EC, Silcox MT, Gunnell GF, Gilbert CC, Yapuncich GS, Allen KL, Welch E, Bloch JI, Gonzalez LA, Kay RF, Seiffert ER. Internal carotid arterial canal size and scaling in Euarchonta: Re-assessing implications for arterial patency and phylogenetic relationships in early fossil primates. Journal of Human Evolution 97:123-144.
Primates differ from other mammals in having bony canals that enclose the branches of the internal carotid artery (ICA) as they pass through the middle ear. The presence and relative size of these canals varies among major primate clades. As a result, differences in the anatomy of the canals for the promontorial and stapedial branches of the ICA have been cited as evidence of haplorhine or strepsirrhine affinities among early fossil euprimates. Here we use microCT to compile the largest quantitative dataset on ICA canal sizes. The data suggest greater variation of the ICA canals within some groups than has been previously appreciated. In species where the promontorial artery supplies the brain with blood, the size of the promontorial canal is significantly correlated with endocranial volume (ECV). In species with alternate routes of encephalic blood supply, the promontorial canal is highly reduced relative to ECV, and correlated with both ECV and cranium size. Ancestral state reconstructions incorporating data from fossils suggest that the last common ancestor of primates had promontorial and stapedial canals that were similar in size and large relative to ECV. This condition is exhibited by treeshrews and most early fossil euprimates, while extant primates exhibit reduction in one canal or another.
Boyer DM, Kirk EC, Silcox MT, Gunnell GF, Gilbert CC, Yapuncich GS, Allen KL, Welch E, Bloch JI, Gonzalez LA, Kay RF, Seiffert ER. Internal carotid arterial canal size and scaling in Euarchonta: Re-assessing implications for arterial patency and phylogenetic relationships in early fossil primates. Journal of Human Evolution 97:123-144.
| Boyer et al 2016 - Internal carotid artery.pdf |
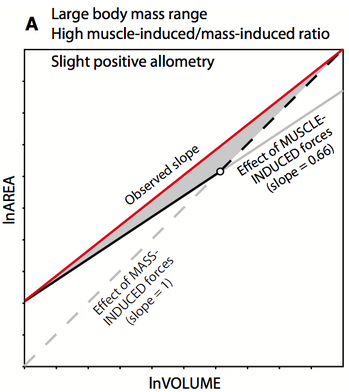
Joint construction in the euarchontan ankle
The articular facets of interosseous joints must transmit forces while maintaining relatively low stresses. To prevent overloading, joints that transmit higher forces should therefore have larger facet areas. The relative contributions of body mass and muscle-induced forces to joint stress are unclear, but generate opposing hypotheses. If mass-induced forces dominate, facet area should scale with positive allometry to body mass. Alternatively, muscle-induced forces should cause facets to scale isometrically with body mass. To re-assess these scaling relationships, we examine the relationship between body size and articular surface areas of the talus. We also examine the scaling pattern of hindlimb muscle PCSA to talar articular surface area, a direct test of the effect of mass-induced forces on joint surfaces. Among most groups, there is an overall trend toward positive allometry for articular surfaces. PCSA generally scales with negative allometry to articular surface area, which indicates joint surfaces increase faster than muscles’ ability to generate force. We suggest a synthetic model to explain the complex patterns observed for talar articular surface area scaling: whether ‘muscles or mass’ drive articular facet scaling is probably dependent on the body size range of the sample and the role of the facet. The relationship between ‘muscle vs. mass’ dominance is likely bone- and facet-specific, meaning that some facets should respond primarily to stresses induced by larger body mass, whereas others primarily reflect muscle forces.
Yapuncich GS, Boyer DM. 2014. Interspecific scaling patterns of talar articular surfaces within primates and their closest living relatives. Journal of Anatomy 224:150-172.
The articular facets of interosseous joints must transmit forces while maintaining relatively low stresses. To prevent overloading, joints that transmit higher forces should therefore have larger facet areas. The relative contributions of body mass and muscle-induced forces to joint stress are unclear, but generate opposing hypotheses. If mass-induced forces dominate, facet area should scale with positive allometry to body mass. Alternatively, muscle-induced forces should cause facets to scale isometrically with body mass. To re-assess these scaling relationships, we examine the relationship between body size and articular surface areas of the talus. We also examine the scaling pattern of hindlimb muscle PCSA to talar articular surface area, a direct test of the effect of mass-induced forces on joint surfaces. Among most groups, there is an overall trend toward positive allometry for articular surfaces. PCSA generally scales with negative allometry to articular surface area, which indicates joint surfaces increase faster than muscles’ ability to generate force. We suggest a synthetic model to explain the complex patterns observed for talar articular surface area scaling: whether ‘muscles or mass’ drive articular facet scaling is probably dependent on the body size range of the sample and the role of the facet. The relationship between ‘muscle vs. mass’ dominance is likely bone- and facet-specific, meaning that some facets should respond primarily to stresses induced by larger body mass, whereas others primarily reflect muscle forces.
Yapuncich GS, Boyer DM. 2014. Interspecific scaling patterns of talar articular surfaces within primates and their closest living relatives. Journal of Anatomy 224:150-172.
| Yapuncich and Boyer 2014 - Interspecific scaling patterns of talar articular surfaces within primates and their closest living relatives |
BODY MASS PREDICTION
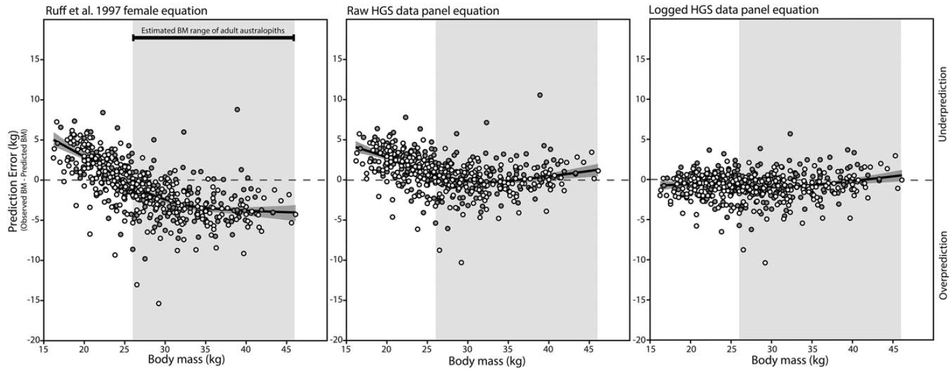
Morphometric panel regression equations for predicting body mass in immature humans
Predicting body mass is a frequent objective of several anthropological subdisciplines, but there are few published methods for predicting body mass in immature humans. Because most reference samples are composed of adults, predicting body mass outside the range of adults requires extrapolation, which may reduce the accuracy of predictions. Prediction equations developed from a sample of immature humans would reduce extrapolation for application to small-bodied target individuals, and should have utility in multiple predictive contexts. Here, we present two novel body mass prediction equations derived from 3468 observations of stature and bi-iliac breadth from a large sample of immature humans (n=173) collected in the Harpenden Growth Study. Prediction equations were generated using raw and natural log-transformed data and modeled using panel regression, which accounts for serial autocorrelation of longitudinal observations. Predictive accuracy was gauged with a global sample of human juveniles (n=530 age- and sex-specific annual means) and compared to the performance of the adult morphometric prediction equation previously identified as most accurate for human juveniles. While the raw data panel equation is only slightly more accurate than the adult equation, the logged data panel equation generates very accurate body mass predictions across both sexes and all age classes of the test sample (mean absolute percentage prediction error = 2.47). The logged data panel equation should prove useful in archaeological, forensic, and paleontological contexts when predictor variables can be measured with confidence and are outside the range of modern adult humans.
Yapuncich GS, Churchill SE, Cameron N, Walker CS. 2018. Morphometric panel regression equations for predicting body mass in immature humans. American Journal of Physical Anthropology. doi: 10.1002/ajpa.23422
Predicting body mass is a frequent objective of several anthropological subdisciplines, but there are few published methods for predicting body mass in immature humans. Because most reference samples are composed of adults, predicting body mass outside the range of adults requires extrapolation, which may reduce the accuracy of predictions. Prediction equations developed from a sample of immature humans would reduce extrapolation for application to small-bodied target individuals, and should have utility in multiple predictive contexts. Here, we present two novel body mass prediction equations derived from 3468 observations of stature and bi-iliac breadth from a large sample of immature humans (n=173) collected in the Harpenden Growth Study. Prediction equations were generated using raw and natural log-transformed data and modeled using panel regression, which accounts for serial autocorrelation of longitudinal observations. Predictive accuracy was gauged with a global sample of human juveniles (n=530 age- and sex-specific annual means) and compared to the performance of the adult morphometric prediction equation previously identified as most accurate for human juveniles. While the raw data panel equation is only slightly more accurate than the adult equation, the logged data panel equation generates very accurate body mass predictions across both sexes and all age classes of the test sample (mean absolute percentage prediction error = 2.47). The logged data panel equation should prove useful in archaeological, forensic, and paleontological contexts when predictor variables can be measured with confidence and are outside the range of modern adult humans.
Yapuncich GS, Churchill SE, Cameron N, Walker CS. 2018. Morphometric panel regression equations for predicting body mass in immature humans. American Journal of Physical Anthropology. doi: 10.1002/ajpa.23422
| Yapuncich et al. 2018 - Morphometric panel regression equations for predicting body mass.pdf |
Alternative methods for calculating percentage prediction error
Since body mass covaries with many ecological aspects of an animal, body mass prediction of fossil taxa is a frequent goal of paleontologists. Body mass prediction often relies on a body mass prediction equation (BMPE): a bivariate relationship between a predictor variable (e.g., molar occlusal area, femoral head breadth) and body mass as observed in extant taxa. A variety of metrics have been used to assess the reliability of BMPEs, including percentage prediction error (%PE), which involves predicting body masses of a test sample comprised of individuals with associated masses. A mean %PE can be calculated in two ways: 1) as the mean %PE of multiple individual predictions (%MPE), or 2) as the %PE of mean body mass generated from the mean predictor value of multiple individuals (here termed %PEM). Differences between these two approaches have never been formally examined and no formal protocols have been recommended. Using a large sample of cercopithecoid primates (406 individuals from 50 species/subspecies) with associated body masses, body mass is predicted with six previously published interspecific BMPEs. Both %MPE and %PEM are calculated and compared. For all BMPEs, the distributions of differences between %MPE and %PEM exhibit positive skew and have medians significantly different from zero, indicating that the examined prediction equations are more accurate at predicting mean body mass than individual body masses. The decreased predictive accuracy of %MPE relative to %PEM likely stems from changing the unit of analysis from mean values (in the reference sample) to individual values (in the test sample) when calculating %MPE. Empirical results are supported with a simulated dataset. Implications for body mass prediction in fossil species are discussed.
Yapuncich GS. 2017. Alternative methods for calculating percentage prediction error and their implications for predicting body mass in fossil taxa. Journal of Human Evolution. doi: 10.1016/j.jhevol.2017.03.001
Since body mass covaries with many ecological aspects of an animal, body mass prediction of fossil taxa is a frequent goal of paleontologists. Body mass prediction often relies on a body mass prediction equation (BMPE): a bivariate relationship between a predictor variable (e.g., molar occlusal area, femoral head breadth) and body mass as observed in extant taxa. A variety of metrics have been used to assess the reliability of BMPEs, including percentage prediction error (%PE), which involves predicting body masses of a test sample comprised of individuals with associated masses. A mean %PE can be calculated in two ways: 1) as the mean %PE of multiple individual predictions (%MPE), or 2) as the %PE of mean body mass generated from the mean predictor value of multiple individuals (here termed %PEM). Differences between these two approaches have never been formally examined and no formal protocols have been recommended. Using a large sample of cercopithecoid primates (406 individuals from 50 species/subspecies) with associated body masses, body mass is predicted with six previously published interspecific BMPEs. Both %MPE and %PEM are calculated and compared. For all BMPEs, the distributions of differences between %MPE and %PEM exhibit positive skew and have medians significantly different from zero, indicating that the examined prediction equations are more accurate at predicting mean body mass than individual body masses. The decreased predictive accuracy of %MPE relative to %PEM likely stems from changing the unit of analysis from mean values (in the reference sample) to individual values (in the test sample) when calculating %MPE. Empirical results are supported with a simulated dataset. Implications for body mass prediction in fossil species are discussed.
Yapuncich GS. 2017. Alternative methods for calculating percentage prediction error and their implications for predicting body mass in fossil taxa. Journal of Human Evolution. doi: 10.1016/j.jhevol.2017.03.001
| Yapuncich 2017 - Alternative methods for calculating percentage prediction error.pdf |
Evaluating morphometric body mass prediction equations with a juvenile test sample
Body mass is an ecologically and biomechanically important variable in the study of hominin biology. Regression equations derived from recent human samples allow for the reasonable prediction of body mass of later, more human-like, and generally larger hominins from hip joint dimensions, but potential
differences in hip biomechanics across hominin taxa render their use questionable with some earlier taxa (i.e., Australopithecus spp.). Morphometric prediction equations using stature and bi-iliac breadth avoid this problem, but their applicability to early hominins, some of which differ in both size and
proportions from modern adult humans, has not been demonstrated. Here we use mean stature, bi-iliac breadth, and body mass from a global sample of human juveniles ranging in age from 6 to 12 years (n = 530 age- and sex-specific group annual means from 33 countries/regions) to evaluate the accuracy
of several published morphometric prediction equations when applied to small humans. Though the body proportions of modern human juveniles likely differ from those of small-bodied early hominins, human juveniles (like fossil hominins) often differ in size and proportions from adult human reference
samples and, accordingly, serve as a useful model for assessing the robustness of morphometric prediction equations. Morphometric equations based on adults systematically underpredict body mass in the youngest age groups and moderately overpredict body mass in the older groups, which fall in the
body size range of adult Australopithecus (26-46 kg). Differences in body proportions, notably the ratio of lower limb length to stature, influence predictive accuracy. Ontogenetic changes in these body proportions likely influence the shift in prediction error (from under- to overprediction). However, because morphometric equations are reasonably accurate when applied to this juvenile test sample, we argue these equations may be used to predict body mass in small-bodied hominins, despite the potential for some error induced by differing body proportions and/or extrapolation beyond the original reference
sample range.
Walker CS, Yapuncich GS (co-authors), Sridhar S, Cameron N, Churchill SE. 2017. Evaluating morphometric body mass prediction equations with a juvenile test sample: accuracy and applicability to small-bodied hominins. Journal of Human Evolution. doi: 10.1016/j.jhevol.2017.03.009
Body mass is an ecologically and biomechanically important variable in the study of hominin biology. Regression equations derived from recent human samples allow for the reasonable prediction of body mass of later, more human-like, and generally larger hominins from hip joint dimensions, but potential
differences in hip biomechanics across hominin taxa render their use questionable with some earlier taxa (i.e., Australopithecus spp.). Morphometric prediction equations using stature and bi-iliac breadth avoid this problem, but their applicability to early hominins, some of which differ in both size and
proportions from modern adult humans, has not been demonstrated. Here we use mean stature, bi-iliac breadth, and body mass from a global sample of human juveniles ranging in age from 6 to 12 years (n = 530 age- and sex-specific group annual means from 33 countries/regions) to evaluate the accuracy
of several published morphometric prediction equations when applied to small humans. Though the body proportions of modern human juveniles likely differ from those of small-bodied early hominins, human juveniles (like fossil hominins) often differ in size and proportions from adult human reference
samples and, accordingly, serve as a useful model for assessing the robustness of morphometric prediction equations. Morphometric equations based on adults systematically underpredict body mass in the youngest age groups and moderately overpredict body mass in the older groups, which fall in the
body size range of adult Australopithecus (26-46 kg). Differences in body proportions, notably the ratio of lower limb length to stature, influence predictive accuracy. Ontogenetic changes in these body proportions likely influence the shift in prediction error (from under- to overprediction). However, because morphometric equations are reasonably accurate when applied to this juvenile test sample, we argue these equations may be used to predict body mass in small-bodied hominins, despite the potential for some error induced by differing body proportions and/or extrapolation beyond the original reference
sample range.
Walker CS, Yapuncich GS (co-authors), Sridhar S, Cameron N, Churchill SE. 2017. Evaluating morphometric body mass prediction equations with a juvenile test sample: accuracy and applicability to small-bodied hominins. Journal of Human Evolution. doi: 10.1016/j.jhevol.2017.03.009
| Walker et al. 2017 - Evaluating morphometric body mass prediction equations with a juvenile human test sample.pdf |

Body mass prediction from tarsal articular surfaces
A species’ body mass is a fundamental aspect of its biology, with wide-reaching impacts on life history, diet, and locomotion. Because body mass cannot be directly measured in extinct taxa, reliable body mass predictors are needed to open valuable avenues for understanding their ecology and evolution. Many published body mass prediction equations rely on dental dimensions, but certain skeletal dimensions should have a more direct and consistent relationship with body mass. We present ordinary least squares and multiple regression equations for predicting euarchontan body mass based on measurements of the articular facets of the astragalus and calcaneus. Separate regressions are given for different levels of taxonomic inclusivity, and the reliability of each equation is evaluated with the coefficient of determination, standard error of the estimate, mean prediction error, and the PRESS statistic. We compare prediction errors to published prediction equations that utilize dental and/or tarsal measures. Finally, we examine the effects of taxonomically specific regressions and apply our equations to a diverse set of non-primates. Our results reveal that predictions based on facet areas are more reliable than most linear dental or tarsal predictors. Multivariate approaches are often better than univariate methods, but require more information (making them less useful for fragmentary fossils). While some taxonomically specific regressions improve predictive ability, this is not true for all primate groups. Of the individual facets, the ectal and fibular facets of the astragalus and the calcaneal cuboid facet are the best body mass predictors.
Yapuncich GS, Gladman JT, Boyer DM. 2015. Predicting euarchontan body mass: A comparison of tarsal and dental variables. American Journal of Physical Anthropology DOI: 10.1002/ajpa.22735.
A species’ body mass is a fundamental aspect of its biology, with wide-reaching impacts on life history, diet, and locomotion. Because body mass cannot be directly measured in extinct taxa, reliable body mass predictors are needed to open valuable avenues for understanding their ecology and evolution. Many published body mass prediction equations rely on dental dimensions, but certain skeletal dimensions should have a more direct and consistent relationship with body mass. We present ordinary least squares and multiple regression equations for predicting euarchontan body mass based on measurements of the articular facets of the astragalus and calcaneus. Separate regressions are given for different levels of taxonomic inclusivity, and the reliability of each equation is evaluated with the coefficient of determination, standard error of the estimate, mean prediction error, and the PRESS statistic. We compare prediction errors to published prediction equations that utilize dental and/or tarsal measures. Finally, we examine the effects of taxonomically specific regressions and apply our equations to a diverse set of non-primates. Our results reveal that predictions based on facet areas are more reliable than most linear dental or tarsal predictors. Multivariate approaches are often better than univariate methods, but require more information (making them less useful for fragmentary fossils). While some taxonomically specific regressions improve predictive ability, this is not true for all primate groups. Of the individual facets, the ectal and fibular facets of the astragalus and the calcaneal cuboid facet are the best body mass predictors.
Yapuncich GS, Gladman JT, Boyer DM. 2015. Predicting euarchontan body mass: A comparison of tarsal and dental variables. American Journal of Physical Anthropology DOI: 10.1002/ajpa.22735.
| Yapuncich et al. 2015 - Predicting euarchontan body mass: a comparison of tarsal and dental variables.pdf |
ANATOMY and FUNCTIONAL MORPHOLOGY
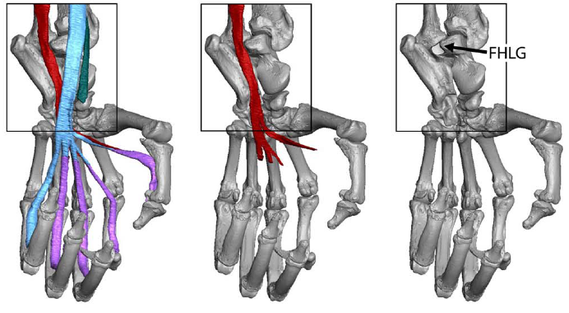
Evolution and behavioral implications of variation in the position and depth of the flexor hallucis longus groove in primates
On the talus, the position and depth of the groove for the flexor hallucis longus tendon have been used to infer phylogenetic affinities and positional behaviors of fossil primates. This study quantifies aspects of the flexor hallucis longus groove (FHLG) to test if: 1) a lateral FHLG is a derived strepsirrhine feature, 2) a lateral FHLG reflects inverted and abducted foot postures, and 3) a deeper FHLG indicates a larger muscle. We used linear measurements of microCT-generated models from a sample of euarchontans (n = 378 specimens, 125 species) to quantify FHLG position and depth. Data are analyzed with ANOVA, Ordinary and Phylogenetic Generalized Least Squares, and Bayesian Ancestral State Reconstruction (ASR). Extant strepsirrhines, adapiforms, plesiadapiforms, dermopterans, and Ptilocercus exhibit lateral FHLGs. Extant anthropoids, subfossil lemurs, and Tupaia have medial FHLGs. FHLGs of omomyiforms and basal fossil anthropoids are intermediate between those of strepsirrhines and extant anthropoids. FHLG position has few correlations with pedal inversion features. Relative FHLG depth is not significantly correlated with body mass. ASRs support a directional model for FHLG position and a random walk model for FHLG depth. The prevalence of lateral FHLGs in many non-euprimates suggest a lateral FHLG is not a derived strepsirrhine feature. The lack of correlations with pedal inversion features suggest a lateral FHLG is not a sufficient indicator of strepsirrhine-like foot postures. Instead, a lateral FHLG may reduce the risk of tendon displacement in abducted foot postures on large diameter supports. A deep FHLG does not indicate a larger muscle, but likely reduces bowstringing during plantarflexion.
Yapuncich GS, Seiffert ER, Boyer DM. 2017. Quantification of the position and depth of the flexor hallucis longus groove in euarchontans, with implications for the evolution of primate positional behavior. American Journal of Physical Anthropology. doi: 10.1002/ajpa.23213
| Yapuncich et al 2017 - Quantification of the position and depth of the flexor hallucis longus groove |
Hallucal metatarsal morphology of Miocene catarrhines from Songhor, Kenya
Songhor is an early Miocene fossil locality in Kenya known for its diverse primate assemblage that includes catarrhine species belonging to the genera Kalepithecus, Limnopithecus, Dendropithecus, Rangwapithecus, and Proconsul. Expeditions to Songhor since the 1930s have recovered unassociated catarrhine postcranial remains from both the fore- and hindlimbs, including multiple elements from the feet. In this study, we describe KNM-SO 31233, a complete left hallucal metatarsal (Mt1), along with several other fragmentary Mt1 specimens (KNM-SO 1080, 5129, 5141, 22235). These fossils were compared to extant catarrhines and platyrrhines, as well as available fossil Miocene catarrhine Mt1s. Morphometric data were obtained from 3D surface renderings and subjected to a number of analyses to assess their phenetic affinity with the comparative sample, make predictions of body mass, and to infer their functional morphology. The size and shape of the Songhor Mt1s are diverse, exhibiting a large robust morph (KNM-SO 5141) similar in size but not in shape to extant African apes, medium-sized morphs (KNM-SO 1080, 5129 and 22235), and a smaller, slender one (KNM-SO 31233) that has a shape resembling arboreal quadrupedal leaping monkeys and suspensory atelines and hylobatids. KNM-SO 31233 is unlike other known fossil Mt1s, and in general, none of the Songhor Mt1s resembled any single extant anthropoid clade or species. The morpho-functional diversity of Songhor Mt1s is consistent with an extensive morphological and phylogenetic catarrhine diversity in the early part of the Miocene epoch.
Patel BA, Yapuncich GS, Tran C, Nengo IO. 2017. Catarrhine hallucal metatarsals from the early Miocene site of Songhor, Kenya. Journal of Human Evolution 108:176-198.
Songhor is an early Miocene fossil locality in Kenya known for its diverse primate assemblage that includes catarrhine species belonging to the genera Kalepithecus, Limnopithecus, Dendropithecus, Rangwapithecus, and Proconsul. Expeditions to Songhor since the 1930s have recovered unassociated catarrhine postcranial remains from both the fore- and hindlimbs, including multiple elements from the feet. In this study, we describe KNM-SO 31233, a complete left hallucal metatarsal (Mt1), along with several other fragmentary Mt1 specimens (KNM-SO 1080, 5129, 5141, 22235). These fossils were compared to extant catarrhines and platyrrhines, as well as available fossil Miocene catarrhine Mt1s. Morphometric data were obtained from 3D surface renderings and subjected to a number of analyses to assess their phenetic affinity with the comparative sample, make predictions of body mass, and to infer their functional morphology. The size and shape of the Songhor Mt1s are diverse, exhibiting a large robust morph (KNM-SO 5141) similar in size but not in shape to extant African apes, medium-sized morphs (KNM-SO 1080, 5129 and 22235), and a smaller, slender one (KNM-SO 31233) that has a shape resembling arboreal quadrupedal leaping monkeys and suspensory atelines and hylobatids. KNM-SO 31233 is unlike other known fossil Mt1s, and in general, none of the Songhor Mt1s resembled any single extant anthropoid clade or species. The morpho-functional diversity of Songhor Mt1s is consistent with an extensive morphological and phylogenetic catarrhine diversity in the early part of the Miocene epoch.
Patel BA, Yapuncich GS, Tran C, Nengo IO. 2017. Catarrhine hallucal metatarsals from the early Miocene site of Songhor, Kenya. Journal of Human Evolution 108:176-198.
| Patel et al. 2017 - Catarrhine hallucal metatarsals from the early Miocene site of Songhor Kenya.pdf |
Virtual endocasts and brain evolution in adapiform primates
Well-preserved crania of notharctine adapiforms from the Eocene of North America provide the best direct evidence available for inferring neuroanatomy and encephalization in early euprimates. Virtual endocasts of the notharctines Notharctus tenebrosus and Smilodectes gracilis from the middle Eocene Bridger formation of Wyoming, and the late Eocene European adapid adapiform Adapis parisiensis, were reconstructed from microCT data. While the three species share many neuroanatomical similarities differentiating them from plesiadapiforms and extant euprimates, our sample of N. tenebrosus displays more variation than S. gracilis, possibly related to differences in the patterns of cranial sexual dimorphism or within-lineage evolution. Body masses predicted from associated teeth suggest that N. tenebrosus was larger and had a lower encephalization quotient (EQ) than S. gracilis, despite their close relationship and similar inferred ecologies. Meanwhile, body masses predicted from cranial length of the same specimens suggest that the two species were more similar, with overlapping body mass and EQ, although S. gracilis exhibits a range of EQs shifted upwards relative to that of N. tenebrosus. While associated data from other parts of the skeleton are mostly lacking for specimens included in this study, measurements for unassociated postcrania attributed to these species yield body mass and EQ estimates that are also more similar to each other than those based on teeth. Regardless of the body mass prediction method used, results suggest that the average EQ of adapiforms was similar to that of plesiadapiforms, only overlapped the lower quadrant for the range of extant strepsirrhines, and did not overlap with the range of extant haplorhines. However, structural changes evident in these endocasts suggest that early euprimates relied more on vision than olfaction relative to plesiadapiforms, despite having relatively small endocranial volumes compared to extant taxa.
Harrington AR, Silcox MT, Yapuncich GS, Boyer DM, Bloch JI. 2016. First virtual endocasts of adapiform primates. Journal of Human Evolution DOI:10.1016/j.jhevol.2016.06.005
Well-preserved crania of notharctine adapiforms from the Eocene of North America provide the best direct evidence available for inferring neuroanatomy and encephalization in early euprimates. Virtual endocasts of the notharctines Notharctus tenebrosus and Smilodectes gracilis from the middle Eocene Bridger formation of Wyoming, and the late Eocene European adapid adapiform Adapis parisiensis, were reconstructed from microCT data. While the three species share many neuroanatomical similarities differentiating them from plesiadapiforms and extant euprimates, our sample of N. tenebrosus displays more variation than S. gracilis, possibly related to differences in the patterns of cranial sexual dimorphism or within-lineage evolution. Body masses predicted from associated teeth suggest that N. tenebrosus was larger and had a lower encephalization quotient (EQ) than S. gracilis, despite their close relationship and similar inferred ecologies. Meanwhile, body masses predicted from cranial length of the same specimens suggest that the two species were more similar, with overlapping body mass and EQ, although S. gracilis exhibits a range of EQs shifted upwards relative to that of N. tenebrosus. While associated data from other parts of the skeleton are mostly lacking for specimens included in this study, measurements for unassociated postcrania attributed to these species yield body mass and EQ estimates that are also more similar to each other than those based on teeth. Regardless of the body mass prediction method used, results suggest that the average EQ of adapiforms was similar to that of plesiadapiforms, only overlapped the lower quadrant for the range of extant strepsirrhines, and did not overlap with the range of extant haplorhines. However, structural changes evident in these endocasts suggest that early euprimates relied more on vision than olfaction relative to plesiadapiforms, despite having relatively small endocranial volumes compared to extant taxa.
Harrington AR, Silcox MT, Yapuncich GS, Boyer DM, Bloch JI. 2016. First virtual endocasts of adapiform primates. Journal of Human Evolution DOI:10.1016/j.jhevol.2016.06.005
| Harrington et al 2016 - First virtual endocasts of adapiforms |
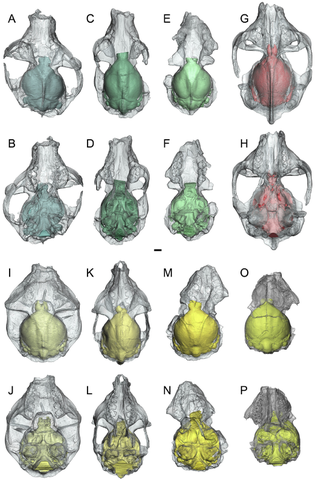
Evolution and implications of medial tibial facet variation in primates
A relatively small medial tibial facet (MTF) of the talus (=astragalus) has been proposed as a synapomorphy uniting anthropoid primates. On the other hand, the distinctive form of the MTF in non-anthropoid primates has been interpreted as a correlate of “prosimian” locomotor tendencies that emphasize vertical climbing and use of inverted, abducted foot postures on small diameter supports, as well as a potential synapomorphy distinguishing non-anthropoid haplorhines and strepsirrhines from non-euprimate euarchontans. Comprehensive quantification of this morphology has been lacking. Here we use scale digital models of tali (n=378 specimens, 122 species) to compute three types of variables that capture different aspects of MTF form: 1) MTF area relative to body mass and ectal facet area; 2) MTF shape (circular vs. elliptical); and 3) dorsal restriction of the MTF on the talar body (i.e., extensive vs. minimal exposure of non-articular area). Results confirm that anthropoids and “prosimians” are generally distinguished from one another. MTF area (but not shape or dorsal restriction) correlates with fibular facet angle of the talus, which has also been argued to reflect inverted foot postures. These results support a functional-adaptive basis for variation in MTF proportions. Ancestral state reconstruction at basal nodes in primate phylogeny reveal little change in the MTF along the primate stem. Basal haplorhines and strepsirrhines independently evolved relatively large and dorsoplantarly deep MTFs, while anthropoids re-evolved morphologies similar to non-euprimates.
Boyer DM, Yapuncich GS, Butler JE, Dunn RH, Seiffert ER. 2015. Evolution of postural diversity in primates as reflected by the size and shape of the medial tibal facet of the talus. American Journal of Physical Anthropology DOI:10.1002/ajpa.22702
| Boyer et al. 2015 - Evolution of postural diversity in primates as reflected by MTF size and shape |
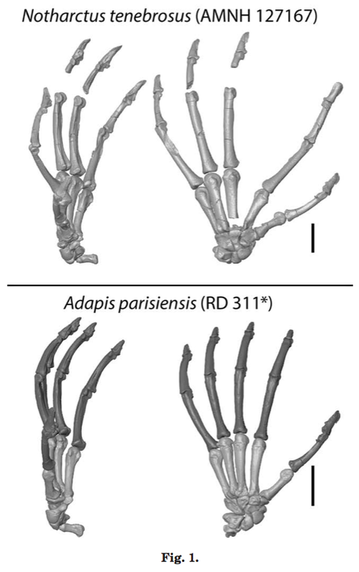
Hands of Paleogene primates
Questions surrounding the origin and early evolution of primates continue to be the subject of debate. Though anatomy of the skull and inferred dietary shifts are often the focus, detailed studies of postcrania and inferred locomotor capabilities can also provide crucial data that advance understanding of transitions in early primate evolution. In particular, the hand skeleton includes characteristics thought to reflect foraging, locomotion, and posture. Here we review what is known about the early evolution of primate hands from a comparative perspective that incorporates data from the fossil record. Additionally, we provide new comparative data and documentation of skeletal morphology for Paleogene plesiadapiforms, notharctines, cercamoniines, adapines, and omomyiforms. Finally, we discuss implications of these data for understanding locomotor transitions during the origin and early evolutionary history of primates. Known plesiadapiform species cannot be differentiated from extant primates based on either intrinsic hand proportions or hand-to-body size proportions. Nonetheless, the presence of claws and a different metacarpophalangeal joint form in plesiadapiforms indicate different grasping mechanics. Notharctines and cercamoniines have intrinsic hand proportions with extremely elongated proximal phalanges and digit rays relative to metacarpals, resembling tarsiers and galagos. But their hand-to-body size proportions are typical of many extant primates (unlike those of tarsiers, and possibly Teilhardina, which have extremely large hands). Non-adapine adapiforms and omomyids exhibit additional carpal features suggesting more limited dorsiflexion, greater ulnar deviation, and a more habitually divergent pollex than observed plesiadapiforms. Together, features differentiating adapiforms and omomyiforms from plesiadapiforms indicate increased reliance on vertical prehensile-clinging and grasp-leaping, possibly in combination with predatory behaviors in ancestral euprimates.
Boyer DM, Yapuncich GS, Chester SGB, Bloch JI, Godinot M. 2013. Hands of early primates. Yearbook of Physical Anthropology 152:33-78.
| Boyer et al. 2013 - Hands of early primates |
SHAPE ANALYSIS
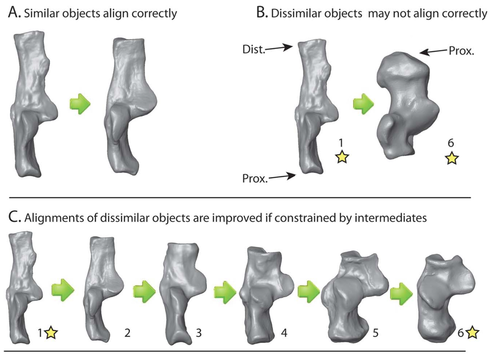
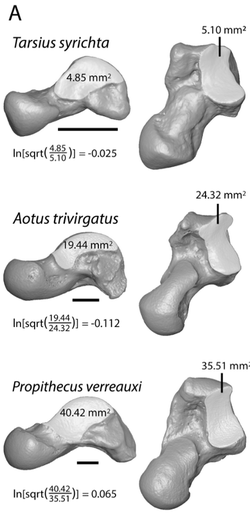
Automatic 3D geometric morphometrics
Three-dimensional geometric morphometric (3DGM) methods have become increasingly sophisticated in the last 20 years, including greater degrees of automation. One aspect shared by all 3DGM methods is that the researcher must designate initial landmarks. Thus, researcher interpretations of homology and correspondence influence shape representation. We present an algorithm allowing fully automatic placement of correspondence points on samples of 3D digital models, which can then be input into standard 3DGM software and analyzed with dimension reduction techniques. Our analyses returned strong correlations between principal component scores, similar variance partitioning among components, and similarities between the shape spaces generated by the automatic and traditional methods. Overall these results suggest that automatic quantifications can lead to shape spaces that are as meaningful as those based on observer landmarks, potentially saving time in data collection, increasing morphological quantification, eliminating observer error, and allowing comparisons of shape diversity between different types of bones. We provide an R package for implementing this analysis.
Boyer DM, Puente J, Gladman JT, Glynn C, Mukherjee S, Yapuncich GS, Daubechies I. 2015. A new fully automated approach for aligning and comparing shapes. The Anatomical Record 298:249-276.
Three-dimensional geometric morphometric (3DGM) methods have become increasingly sophisticated in the last 20 years, including greater degrees of automation. One aspect shared by all 3DGM methods is that the researcher must designate initial landmarks. Thus, researcher interpretations of homology and correspondence influence shape representation. We present an algorithm allowing fully automatic placement of correspondence points on samples of 3D digital models, which can then be input into standard 3DGM software and analyzed with dimension reduction techniques. Our analyses returned strong correlations between principal component scores, similar variance partitioning among components, and similarities between the shape spaces generated by the automatic and traditional methods. Overall these results suggest that automatic quantifications can lead to shape spaces that are as meaningful as those based on observer landmarks, potentially saving time in data collection, increasing morphological quantification, eliminating observer error, and allowing comparisons of shape diversity between different types of bones. We provide an R package for implementing this analysis.
Boyer DM, Puente J, Gladman JT, Glynn C, Mukherjee S, Yapuncich GS, Daubechies I. 2015. A new fully automated approach for aligning and comparing shapes. The Anatomical Record 298:249-276.
| Boyer et al. 2015 - A new fully automated approach for aligning and comparing shapes |
PHYLOGENETIC COMPARATIVE METHODS
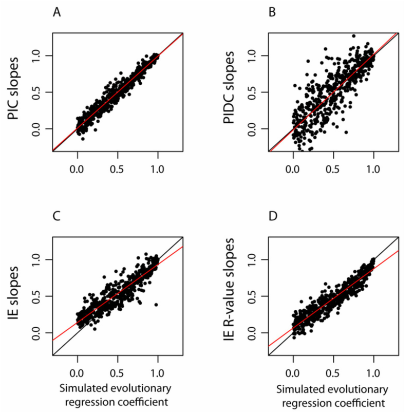
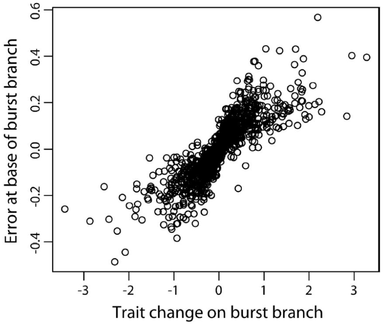
Evaluation of the Independent Evolution method
Phylogenetic comparative methods (PCMs) use data on species traits and phylogenetic relationships to shed light on evolutionary questions. Recently, Smaers and Vinicius (2009) suggested a new PCM, Independent Evolution (IE), which purportedly employs a novel model of evolution based on Felsenstein’s (1988) Adaptive Peak Model. The authors found that IE improves upon previous PCMs by producing more accurate estimates of ancestral states, as well as separate estimates of evolutionary rates for each branch in a phylogeny. Here, we document substantial theoretical and computational issues with IE. When data are simulated under Brownian motion, IE produces severely biased estimates of ancestral states and changes along individual branches. We show that these branch-specific changes are essentially ancestor-descendant or “directional” contrasts (sensu Harvey and Pagel, 1991), and draw parallels between IE and previous PCMs such as “minimum evolution” (Huey and Bennett, 1987). Additionally, while comparisons of branch-specific changes between variables have been interpreted as reflecting the relative strength of selection on those traits, we demonstrate that regressing IE estimated branch-specific changes against one another gives a biased estimate of the scaling relationship between these variables, and provides no advantages beyond established PCMs . In light of our findings, we discuss the results of previous papers that employed IE. We conclude that IE is not a viable PCM, and should not be used in comparative analyses.
RH Griffin, Yapuncich GS. 2015. The Independent Evolution method is not a viable phylogenetic comparative method. PLoS ONE 10(12): e0144147. doi:10.1371/journal.pone.0144147
| Griffin and Yapuncich 2015 - IE is not a viable PCM |
A critical comment on the 'multiple variance Brownian motion' model of Smaers et al. (2016)
Smaers, Mongle & Kandler (2016) (Biological Journal of the Linnean Society, 118: 78-94) introduced a new phylogenetic comparative method, multiple variance Brownian motion (mvBM), for reconstructing ancestral states given a phylogenetic tree and continuous trait data. The authors conducted a simulation study and argued that mvBM outperforms constant variance Brownian motion (BM) when rates of evolution vary across the phylogeny. In this comment, we argue that mvBM is not a viable statistical method because it is fundamentally a circular analysis that overfits phylogenetic branch lengths to the data. We further argue that the comparison of mvBM to BM under conditions where the assumptions of BM are clearly violated is not an informative performance analysis, and that the simulation study of Smaers et al. (2016) exaggerates the performance of mvBM by focusing on a narrow range of simulation conditions and reporting aggregated accuracy metrics that obscure severe inaccuracy and bias in its ancestral state estimates. Our arguments are supported by simulation results. We conclude that mvBM is not a viable phylogenetic comparative method.
Smaers, Mongle & Kandler (2016) (Biological Journal of the Linnean Society, 118: 78-94) introduced a new phylogenetic comparative method, multiple variance Brownian motion (mvBM), for reconstructing ancestral states given a phylogenetic tree and continuous trait data. The authors conducted a simulation study and argued that mvBM outperforms constant variance Brownian motion (BM) when rates of evolution vary across the phylogeny. In this comment, we argue that mvBM is not a viable statistical method because it is fundamentally a circular analysis that overfits phylogenetic branch lengths to the data. We further argue that the comparison of mvBM to BM under conditions where the assumptions of BM are clearly violated is not an informative performance analysis, and that the simulation study of Smaers et al. (2016) exaggerates the performance of mvBM by focusing on a narrow range of simulation conditions and reporting aggregated accuracy metrics that obscure severe inaccuracy and bias in its ancestral state estimates. Our arguments are supported by simulation results. We conclude that mvBM is not a viable phylogenetic comparative method.
| Griffin and Yapuncich 2016 - Comment on Smaers et al 2016 |